干酪乳杆菌发酵及热加工方式对小麦蛋白抗原性的影响(二)
2 結果与剖析
2.1 干酪乳杆菌的分离出来评定
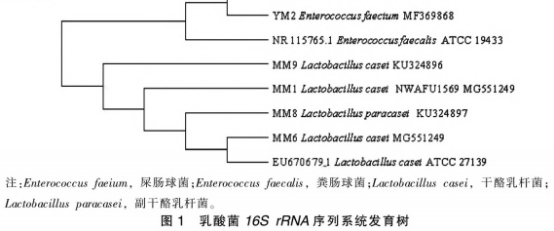
从培养液中任意选择乳白色、网形的突起菌体,经双氧水酶和革兰氏染色实验后,挑选出过氧化氢酶呈阴性、革兰氏阳性菌转录组测序,经同宗序列比对后,共评定出4株干酪乳杆菌和2株粪肠球菌。由图1的系统发育进化树能够看得出,共监测到2个属:肠球菌属和乳酸杆菌属。MMl(干酪乳杆菌)、MM6(干酪乳杆菌)、MM8(副干酪乳杆菌)、MM9(干酪乳杆菌)和YLl(屎肠球菌)、YM2(屎肠球菌)产生了2个种群并与相应的方式菌种聚在一起,进一步认证了分子结构学评定结论的稳定性。
2.2 菌种生长曲线和菌体数与OD值对应关系的测量
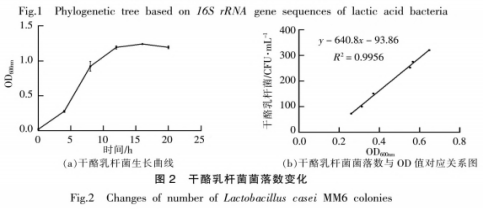
挑选与方式菌种亲缘关系近期的MM6菌种开展发醇实验。如图所示2所显示,干酪乳杆菌MM6在2~14h处在指数值成长期,在14~20h进到生长发育稳定型,20h之后因为营养元素不够和类化合物的累积,菌体身亡总数增加。实验搜集指数值生长发育后期菌体开展发醇实验,这时菌种生长发育充沛,酶等新陈代谢有关化学物质活力较高。根据倒平板记取菌体数,获得其与0D600nm值的对应关系(图2b),纵轴为稀释液106后的菌体数,双蒸水重飘浮菌泥至相对应0D600nm值以确保发酵面团中活菌体的成分为108CFU/g。
2.3 发醇全过程中pH值、TTA值的转变
如图所示3所显示,伴随着发酵时间增加,空缺组(K)pH值和TTA值转变并不大,殊不知干酪乳杆菌加上组(LAB)的pH值逐步减少,24h后低至3.9,TTA值与pH值转变呈相对的发展趋势。干酪乳杆菌归属于异形发醇乳酸菌饮料,发醇全过程中乳酸菌、冰醋酸等有机物的新陈代谢造成 面糊pH值降低、TTA值提升。

2.4 发醇全过程中蛋白质成分及抗原性的转变
选用SDS-PAGE实验对发醇全过程中面糊蛋白质的变动实现了评定。如图所示4所显示,在发醇24h后,清蛋白、血蛋白、醇溶蛋白质、谷蛋白的成分和成分均有转变 。伴随着发醇的开展,清、血蛋白中25~35,15~25,10~15ku杂带变淡:醇溶蛋白质杂带无显著转变:谷蛋白80ku和40ku上下的杂带发醇后显著变淡。12,15~25,40.88ku地区各自相应着麦子过敏源Tria37.Tria18,Tria36和Tria26,从图4能够看得出,这种地区的蛋白质在发醇24h后均有一定程度上的溶解。
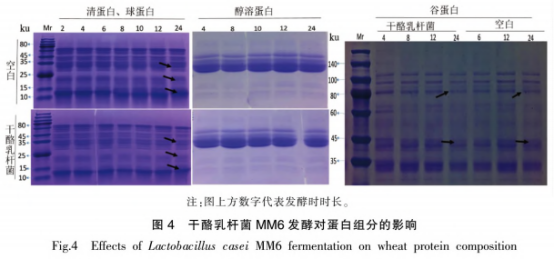
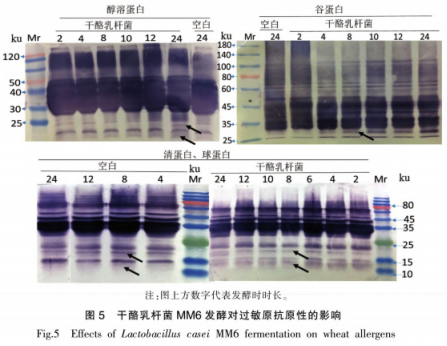
图5表明,发醇后的绝大多数蛋白质试品仍与抗麦子蛋白质兔血清蛋白发生了比较明显的反映。清蛋白和血蛋白中12ku和15~25ku地区的Tria37和Tria18成分大幅度降低,与抗原融合反映伴随着发酵时间的增加而变弱。醇溶蛋白质和谷蛋白在25~30ku地区内慢慢产生了新的免疫力融合杂带,表明一些免疫力肽段可被分解成更小分子质量的蛋白,可是仍具备抗原性,可能是因为过敏源线性表位并未被破环。发醇全过程中面糊蛋白质成分及蛋白质抗原性转变说明,绝大多数麦子过敏源无法根据发醇获得溶解,L0ponen等和Ganzle的分析强调.酸面粉发酵全过程中一部分谷蛋白会被分解成30ku上下的醇溶蛋白质,这与本实验結果相符合。发醇是一种迟缓酶解的全过程,乳酸菌发酵产酸能将面糊的pH值降至4上下,可激话面糊中首要的胰蛋白酶(天冬氨酸胰蛋白酶)溶解生物大分子蛋白质。小麦面粉中的内源性胰蛋白酶及其乳酸菌饮料中的肽酶等均能具有水解反应面糊蛋白质的功效。已经有研究表明干酪乳杆菌具备膜胰蛋白酶,可将管理体系中的生物大分子蛋白质水解反应为活性多肽,膜上的活性多肽装运系统软件会将活性多肽迁移至胞内,胞内肽酶将再次水解反应肽段B181。因而,对比于空缺组,加上干酪乳杆菌的面糊,麦子蛋白质被一部分水解反应。
文章版权备注
- 2023-05-01奶味香精的制备技术与开发现状
- 2023-04-04阿魏酸及其衍生物在食品添加剂领域研究进展(一)
- 2023-03-23甘肃省榆中县市场监督管理局强化食品快检 筑牢食品安全防线
- 2023-03-23常德:专项整治酒类市场12种违法行为
- 2023-03-23贵州这7批次食品检出食品添加剂问题,有食用植物调和油、冰糖大蒜、无油剁椒等
- 2023-03-23怀化:部署开展制止餐饮浪费专项行动
- 2023-03-23超90%展商已确认展位 6月食品原料展带来海内外商机
- 2023-03-23关于召开2023年国际食品安全与健康大会的通知
- 2023-03-23河南省市场监管局召开落实食品安全“两个责任”暨制止餐饮浪费专项行动调度视频会议
- 2023-03-23邵阳市食品安全“两个责任”机制推进暨野生蘑菇中毒防控部署电视电话会议召开
 豫ICP备19024296号
豫ICP备19024296号 售前咨询
售前咨询