两个来自非脱羧勒克菌的MDR质粒耐药机制研究(一)
非脱羧勒克菌(Leclercia adcarboxglata)是一种广泛存在于自然界中的肠杆菌科革兰阴性兼性厌氧菌,与大肠杆菌有许多相同的生化特征,能引起免疫功能不全或有慢性疾病患者的感染,是多种微生物感染的病原体之一,与其他细菌共同引起较为严重的共感染。该菌是一种机会致病菌且与诱发因素有关,如创伤或免疫抑制等,可从血液、粪便、痰、尿、腹膜液和脓液中分离获得。
β‑内酰胺类抗生素在过去70年被广泛作为抗菌药物,其耐药现象可由多种机制引起,如产β‑内酰胺酶、主动外排系统、生物被膜机制、膜孔蛋白缺失以及与青霉素结合蛋白亲和力下降等,其中β‑内酰胺酶水解或修饰β‑内酰胺类抗生素使其失活,使相应菌株对几乎所有β‑内酰胺类抗生素产生耐药性,质粒携带β‑内酰胺耐药基因是导致菌株耐药性快速传播的主要原因。非脱羧勒克菌对头孢菌素、碳青霉烯类、四环素类、氨基糖苷类、喹诺酮类和氯霉素敏感,近年来陆续报道了少量产超广谱β‑内酰胺酶(extended‑spectrumβ‑lactamases,ESBL)和碳青霉烯酶的非脱羧勒克菌感染相关病例。现阶段只有少量多药耐药(multi‑drug resistant,MDR)的非脱羧勒克菌被报道,关于blaCTX‑M的报道仅有2例。
本研究中,我们对来源于原南京军区总医院重症急性胰腺炎患者血液样本的菌株150707804和重庆医科大学附属第一医院胃癌患者胃引流样本的菌株P12375进行了分析,旨在对其遗传背景、耐药表型进行鉴定,通过高通量全基因组序列测定分析,结合生物信息学研究,对其携带耐药基因的移动元件和耐药基因座位进行精细注释,揭示非脱羧勒克菌多重耐药的传播机制。
1 材料和方法
1.1菌株来源与鉴定
菌株150707804分离自47岁男性重症急性胰腺炎患者血液样本,菌株P12375分离自63岁男性胃癌患者引流液样本。16S rDNA基因扩增初步确定菌种。
1.2药敏实验
使用VITEK 2 Compact全自动细菌鉴定及药敏分析仪对菌株150707804、P12375分别进行药敏MIC检测,依据美国临床与实验室标准学会(Clinical and Laboratory Standards Institute,CLSI)2020标准判读结果[20]。
1.3质粒测序及组装
使用Qiagen公司的Ultra Clean®Microbial DNA Isolation kit(CAT.12224‑250)试剂盒,分别从分离株150707804和P12375中提取基因组DNA。构建均值约15 kb(10~20 kb)的DNA文库,通过PacBio RSII测序平台进行“三代”高通量基因组测序。同时,构建均值约400 bp(150~600 bp)的双端测序(paired‑end)文库,利用HiSeq“二代”测序平台上机测序。通过Proovread[21]软件用短双端测序读长(paired‑end reads)校正PacBio长reads,校正过的PacBio reads通过HGAP v3.0(基因组覆盖率100×)进行拼接组装,最终获得野生株完整基因组全序。
1.4生物信息学分析
以复制子repA正向序列起始密码子的第一个碱基作为整个质粒的“+1”位点,首先采用RAST 2.0初步预测质粒的开放阅读框(open reading frame,ORF),结合BLASTP/BLASTN、RefSeq数据库和Conserved Domains数据库进行ORF和假基因的逐个功能分析。利用在线数据库ResFinder和CARD对耐药基因进行注释,ISfinder、INTEGRALL和Tn Number Registry分别对插入序列、整合子、转座子等结构进行注释。用Inkscape 1.0绘制质粒整体结构图、质粒近源结构比较图和耐药基因座位精细结构比较图。
1.5序列登录号
两株菌进行全基因组测序后,得到质粒p707804‑CTXM和pP12375‑CTXM序列,提交至Gen‑Bank,登录号分别为MN823992和MN821366。
2 结果
2.1菌株鉴定及耐药基因筛查
菌株150707804和P12375经16S rDNA扩增初步鉴定,全基因组DNA序列与非脱羧勒克菌USDA‑ARS‑USMARC‑60222(登录号:NZ_CP013990)进行平均核苷酸一致性(average nucleotide identity,ANI)比对,结果菌株150707804为98.66%、菌株P12375为98.72%,最终核定两株菌均为非脱羧勒克菌。
2.2药敏测定结果
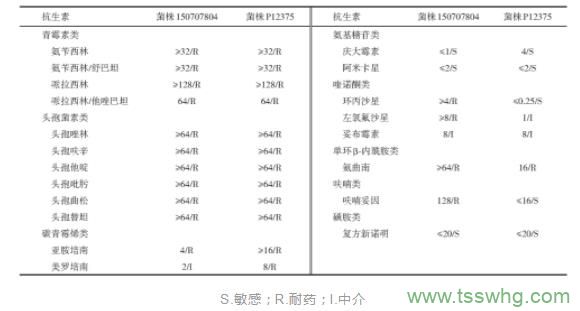
野生株150707804对青霉素类、头孢菌素类、碳青霉烯类、喹诺酮类、单环β‑内酰胺类及呋喃类抗生素耐药,对氨基糖苷类和磺胺类抗生素敏感;野生株P12375对青霉素类、头孢菌素类、碳青霉烯类、单环β‑内酰胺类及呋喃类抗生素耐药,对氨基糖苷类、喹诺酮和磺胺类抗生素敏感(表1),其中,喹诺酮类和呋喃类耐药表型不同,可能原因:(1)喹诺酮类抗生素的主要作用靶点为DNA螺旋酶和拓扑异构酶Ⅳ,使细菌DNA不能形成超螺旋。质粒p707804‑CTXM和pP12375‑CTXM无喹诺酮类耐药基因,其对应染色体上也不含有耐药基因,但菌株150707804和P12375的其他质粒中分别含有qnrA、aac(6′)Ib-cr和qnrS、aac(6')Ib-cr。P12375对喹诺酮类抗生素敏感可能是由于耐药基因突变不表达所致;(2)呋喃类抗生素主要由对氧不敏感的硝基还原酶基因(nfsA和nfsB)突变失活引起,干扰氧化还原酶从而阻断细菌的正常糖代谢。质粒p707804‑CTXM和pP12375‑CTXM不含呋喃类耐药基因,且菌株150707804和P12375的其他质粒及染色体上也不含该类耐药基因,可能是由于外排泵等其他耐药机制引起表型差异,并不属于本研究的范围。
2.3质粒基因组序列测定及生物信息学分析
2.3.1质粒p707804‑CTXM和pP12375‑CTXM的基本特征
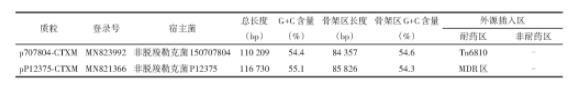
p707804‑CTXM和pP12375‑CTXM大小分别为110.2和116.7 kb(图1,表2)。两个质粒全序高度相似(覆盖度75%、核苷酸同源性98.69%),均包含保守骨架区(覆盖度85%、核苷酸同源性98.69%)和外源插入区。
2.3.2质粒骨架区
质粒p707804‑CTXM和pP12375‑CTXM骨架区总长度分别为84.3和85.8 kb(图2)。二者均由质粒复制相关区(plasmid replication)、质粒稳定相关区(plasmid maintenance)以及质粒接合转移相关区(conjugal transfer)组成。两个质粒有一定保守性,它们含有相同的复制子repA,为同一类型质粒;含相同的质粒稳定相关基因parA、stbAB等;接合转移区均含tra和pil两个基因簇。同时,这两个质粒也存在差异性:(1)pP12375‑CTXM中orf531被MDR区分割为两部分,而在p707804‑CTXM中orf531完整;(2)两质粒orf531至parA间差异性较大,有大片段的序列置换;(3)p707804‑CTXM的orf516与pP12375‑CTXM的orf927存在基因置换;(4)接合转移区内p707804‑CTXM的orf774、orf234与pP12375‑CTXM的orf1890存在基因置换。
声明:本文所用图片、文字来源《师范大学学报》,版权归原作者所有。如涉及作品内容、版权等问题,请与本网联系删除。
相关链接:青霉素,碳青霉烯,氨基糖苷
文章版权备注
- 2023-05-01不同贮藏方式对红香酥梨采后生理及品质的影响(一)
- 2023-05-01奶味香精的制备技术与开发现状
- 2023-05-01双孢蘑菇液体菌种发酵及栽培效果浅析
- 2023-05-01菌种强化结合工艺优化提高酱香白酒基酒中四甲基吡嗪含量的研究(二)
- 2023-05-01发酵小麦胚芽产2,6-二甲氧基对苯醌菌种筛选及发酵条件优化(一)
- 2023-05-01猕猴桃中铁含量的测定与测定因素的探究
- 2023-05-01腐植酸调节砷酸盐生菜毒性作用研究(四)
- 2023-05-01腐植酸调节砷酸盐生菜毒性作用研究(三)
- 2023-05-01腐植酸调节砷酸盐生菜毒性作用研究(二)
- 2023-05-01腐植酸调节砷酸盐生菜毒性作用研究(一)
 豫ICP备19024296号
豫ICP备19024296号 售前咨询
售前咨询