3.2 效用物对α-葡萄糖苷酶的抑止原理
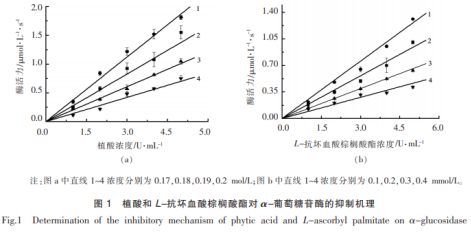
2效用物对α-葡萄糖苷酶的抑止原理植酸和L-抗坏血酸豆蔻酸酯对α-葡萄糖苷酶抑止原理如图所示1所显示。图上的曲线图均给予起点,直线斜率各自随效用物浓度值增高而减少。因而,2种效用物对α-葡萄糖苷酶均为交叉性抑止,效用物与酶以非共价键融合成可离解的一氧化氮合酶。在该酶活管理体系的测量中尽管酶催化反应速度速度减少,可是沒有降低合理酶量。
3.3 效用物对α-葡萄糖苷酶的抑止种类
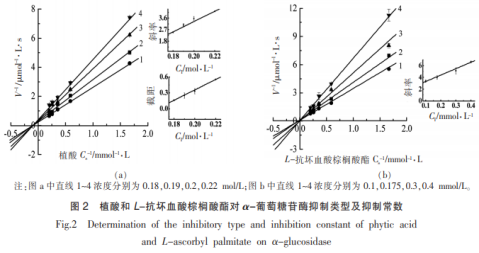
植酸和L-抗坏血酸豆蔻酸酯对α-葡萄糖苷酶抑止种类如图2所显示。图2a所显示,植酸的Lineweaver-Burk图中平行线1~4交叉于第Ⅱ象限,伴随着植酸浓度值上升,米氏常数(Km)增大,较大化学反应速率(Vmax)减少,说明效用物植酸对α-葡萄糖苷酶是复合型抑止,具备竞争抑止和非竞争抑止2种方式。植酸既能与分散的酶融合,也可以与酶-底物一氧化氮合酶融合。根据制作Dixon图计算抑止参量KI和KIS各自为KI=0.117mol/L和KIS=0.163mol/L。由KI值低于KIS值,说明化学物质与α-葡萄糖苷酶融合以外,关键产生ESI(效用物-酶-底物)一氧化氮合酶。
L-抗坏血酸豆蔻酸酯对α-葡萄糖苷酶抑止种类如图2b所显示,平行线1~4交叉于第Ⅰ象限,十分贴近Y轴,伴随着L-抗坏血酸豆蔻酸酯浓度值上升,米氏常数(Km)增大,较大化学反应速率(Vmax)不会改变(转变轻度),说明L-抗坏血酸豆蔻酸酯对α-葡萄糖苷酶是市场竞争型抑止,主要是与α-葡萄糖苷酶融合,抑止参量KI=0.202mmol/L。
3.4 分子对接結果
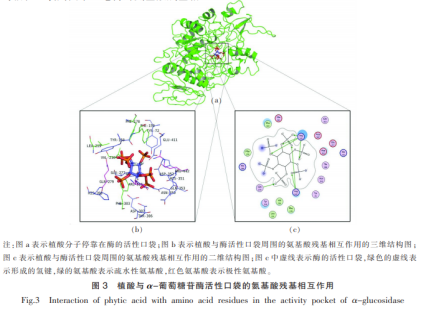
植酸与α-葡萄糖苷酶活力袋子的氨基酸残基相互影响如图所示3所显示。如图所示3a所显示,植酸进到葡萄糖苷酶的活力袋子中,活力袋子周边关键的碳水化合物Phe303,Phe159,Val216,Leu219,Phe178是疏水性碳水化合物,与活力袋子的产生相关,这与植酸进到活力袋子也是有一定的危害。如图所示3b,3c所显示,植酸分子结构含有6个磷酸根离子,所需的阳离子有益于与活力袋子周边带正电的氨基酸残基相互影响。配位分子结构植酸与α-葡萄糖苷酶活力袋子氨基酸残基Arg315,Arg442,Gln279产生共价键。共价键是植酸与α-葡萄糖苷酶融合的主导作用。除此之外,植酸与氨基酸残基Glu411,Gln353,Asp307,Glu277,Asp352,His351,Thr306,His280,Asn350,Tyr72,Tyr158产生范德华力,产生的非共价键相互作用力(共价键和范德华力),在平稳植酸与α-葡萄糖苷酶一氧化氮合酶中起关键功效。
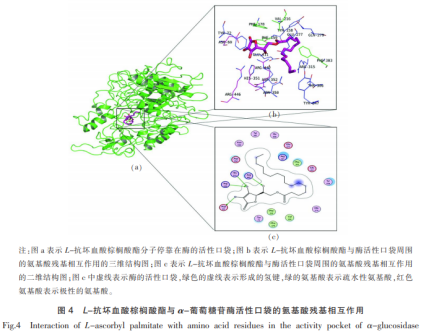
L-抗坏血酸豆蔻酸酯与α-葡萄糖苷酶活力袋子的氨基酸残基相互影响如图4所显示。由图4b,4c能够看得出,L-抗坏血酸豆蔻酸酯的分子式,一端为含有酚羟基的苯环,接着相互连接的是较长碳链构造。含有酚羟基苯环一端进到酶的活性袋子房屋朝向氨基酸残基His315,Arg442,Arg446,Asp69,并与4个氨基酸残基产生共价键。共价键产生的越大,配位分子结构与酶融合越平稳。全部配位分子结构进到酶的活性袋子中,因为碳链较长,进到活力袋子必须扭曲和变化姿态及摆脱室内空间阻拦而耗费大量的动能。除此之外,配位分子结构L-抗坏血酸豆蔻酸酯的尾端房屋朝向Phe303,Phe159,Val216,Phe178疏水性碳水化合物,产生亲水性相互影响。配位分子结构L-抗坏血酸豆蔻酸酯与周边的氨基酸残基Tyr72,Gln279,Tyr158,Asp352,Asn350,Tyr347,Thr306,Glu277,Arg315,Glu411产生范德华力。L-抗坏血酸豆蔻酸酯与α-葡萄糖苷酶的活力袋子周边的碳水化合物十分平稳的融合。
4 结果
抗氧剂植酸和L-抗坏血酸豆蔻酸酯对α葡萄糖苷酶均有控制实际效果。植酸对α-葡萄糖苷酶过半数抑止浓度值为(0.207±3.137)mol/L,抑止种类体现为可逆性复合型抑止。L-抗坏血酸豆蔻酸酯对α葡萄糖苷酶的IC50数值(0.39±0.00838)mmol/L,抑止种类为可逆性市场竞争型抑止。植酸和L-抗坏血酸豆蔻酸酯各自与α-葡萄糖苷酶活力袋子的氨基酸残基相互影响,产生共价键,亲水性相互影响,范德华力等非共价键相互影响。此結果与酶动力学模型的抑止原理的分析效果具备一致性。除此之外,对于2种抗氧剂对酶的控制实际效果,与限制的增加量完成较为,L-抗坏血酸豆蔻酸酯对α-葡萄糖苷酶主要表现出精确性,在做为α-葡萄糖苷酶缓聚剂给予理论来源的与此同时,也为L-抗坏血酸豆蔻酸酯的多功能性运用带来了概念支撑点。
文章版权备注
- 2023-05-01奶味香精的制备技术与开发现状
- 2023-04-04阿魏酸及其衍生物在食品添加剂领域研究进展(一)
- 2023-03-23甘肃省榆中县市场监督管理局强化食品快检 筑牢食品安全防线
- 2023-03-23常德:专项整治酒类市场12种违法行为
- 2023-03-23贵州这7批次食品检出食品添加剂问题,有食用植物调和油、冰糖大蒜、无油剁椒等
- 2023-03-23怀化:部署开展制止餐饮浪费专项行动
- 2023-03-23超90%展商已确认展位 6月食品原料展带来海内外商机
- 2023-03-23关于召开2023年国际食品安全与健康大会的通知
- 2023-03-23河南省市场监管局召开落实食品安全“两个责任”暨制止餐饮浪费专项行动调度视频会议
- 2023-03-23邵阳市食品安全“两个责任”机制推进暨野生蘑菇中毒防控部署电视电话会议召开
 豫ICP备19024296号
豫ICP备19024296号 售前咨询
售前咨询